La obra de Richard Lewontin (1929-2022) entreteje trabajos y contribuciones en campos muy diversos de la biología y la reflexión crítica sobre la actividad científica. Desde mi punto de vista, esas contribuciones forman parte de un esfuerzo por comprender los procesos del mundo vivo y la disciplina que los estudia desde una visión integral de la realidad y de nuestras teorías para entenderla, en lo que el marxismo clásico llamó la totalidad (Lukács, 1985).
Lewontin fue uno de los más claros críticos del determinismo genético y al mismo tiempo un genetista que se mantuvo hasta el final haciendo contribuciones muy importantes en el uso de datos genéticos para inferir procesos evolutivos, como el estudio de la historia demográfica de las poblaciones (véase, por ejemplo, Shpak et al., 2010) y el análisis de las huellas de procesos selectivos y neutrales (véase Garrigan et al., 2010). En su obra es posible ver que las preguntas que se planteó y las herramientas que usó para contestarlas surgen de poner atención en las relaciones entre los componentes de los sistemas biológicos.
El desequilibrio de ligamiento, huella de los procesos evolutivos
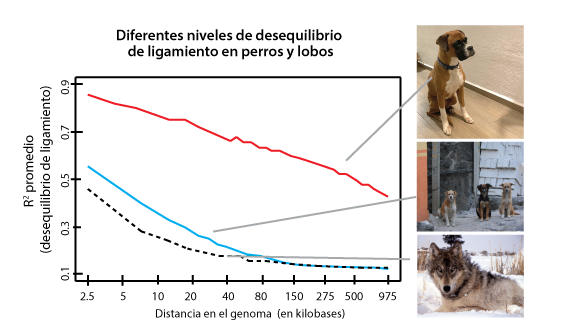
El desequilibrio de ligamiento consiste en la asociación no azarosa o aleatoria de los alelos en dos loci dentro de las gametas. A partir de la segunda ley de Mendel, se esperaría que las diferentes combinaciones de los alelos de dos loci se formarían de manera proporcional a las frecuencias de cada uno de los alelos (véase también Eguiarte y Souza, en esta colección). Sin embargo, en muchos casos esto no sucede, en primera instancia porque los loci (que pueden ser genes o secuencias no codificantes) pueden estar ligados físicamente y son heredados juntos en un mismo cromosoma. A esto hay que añadir que diferentes procesos evolutivos, como la deriva génica, la endogamia o la selección natural, pueden provocar que ciertos haplotipos en las gametas se presenten en frecuencias mayores o menores que las esperadas por azar. A Lewontin le interesaba mucho estudiar este fenómeno. En 1960 propuso junto con Kojima una herramienta estadística, que llamó D, para medir el grado de desequilibrio de ligamiento, es decir, el grado en que los alelos en dos o más loci aparecen asociados con mayor (o menor) frecuencia de lo esperado por azar dadas sus frecuencias individuales. Esta herramienta fue modificada primero por el propio Lewontin en 1964, y luego por Hill y Robertson en 1968, pero la concepción original de Lewontin y Kojima —consistente en mirar, desde la perspectiva de los alelos que la compongan, las frecuencias de los tipos de gametas que se forman en una población— sigue siendo la base de lo que conocemos sobre el desequilibrio de ligamiento.
Conocer los niveles de desequilibrio de ligamiento es importante porque nos permite inferir su efecto en procesos evolutivos del pasado. Por ejemplo, procesos como la consanguinidad o la selección natural pueden manifestarse de manera diferente en la cantidad de desequilibrio de ligamiento que observamos en una población u otra.
En cambio, el tamaño de los bloques de desequilibrio de ligamiento (los grupos de pares de bases que se heredan como paquetes, con nula o muy rara recombinación) ocasionados por la deriva génica está más relacionado con el tamaño poblacional. En el caso de la selección natural, ese tamaño depende, entre otras cosas, de la fuerza de la selección y la tasa de recombinación presente en una especie (r), la cual también altera la forma en como actúan los diferentes procesos evolutivos. Así, el desequilibrio de ligamiento es a la vez causa y efecto de diferentes procesos microevolutivos, de tal forma que abre otra “ventana hacia el pasado”, es decir, es una herramienta para inferir procesos evolutivos que sucedieron en el pasado.
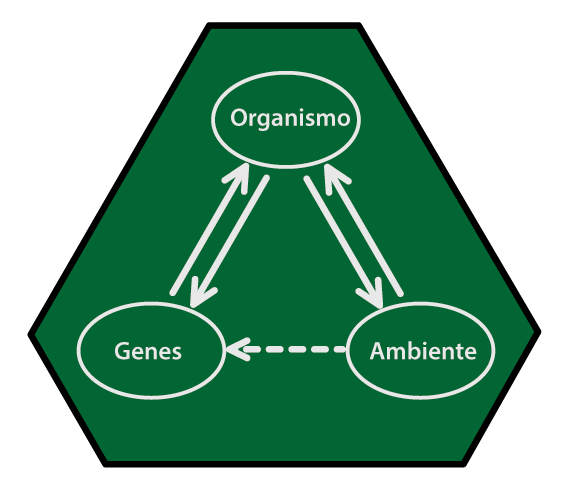
De esta manera, en la obra de Lewontin la relevancia de estudiar la variación genética (véase Eguiarte y Souza en este número) no estriba en pensar que los genes son agentes causales o los elementos más importantes ontológicamente hablando. Más que causas, los genes son un registro del pasado, y contar con registros del pasado es sumamente importante para una ciencia histórica como la biología.
En esto coinciden —entre otras cosas— Lewontin y Stephen Jay Gould (1941-2002). Este último, en una colección de textos editada en 2002 (Singh, Krimbas, Paul y Beatty (eds.) 2002, Thinking about evolution, Cambridge University Press) para honrar a Lewontin, usó la metáfora del genoma como un “libro de registro” en el que quedan asentados diferentes procesos evolutivos —entre los cuales la selección natural es uno más— que operan en diferentes niveles de organización de la materia (organismos, poblaciones, comunidades, etc.) y que dejan sus huellas en el genoma. Los biólogos, entonces, estudiamos los genes como unidad de registro porque en ellos se mantiene una especie de bitácora de cambios derivados de los otros niveles de organización (Gould, 2003).